Morphological characterization of 23 Malus domestica Borkh cultivars from central Spain
Abstract
The purpose of this work was to morphologically characterize an apple tree collection composed of 67 individuals from 41 accessions belonging to 23 old Spanish apple cultivars (Malus domestica Borkh) alongside 9 reference cultivars. The studied germplasm was collected previously in rural areas of central Spain (Sierra Norte de Madrid and Tagus river basin) and it was analyzed through 67 descriptors mainly from IBPGR and UPOV. We found a very high morphological diversity in the studied old apple cultivars, as 48% of the descriptors (most of them devoted to fruit traits) were significantly different between types of cultivars. In addition, the sample cultivars resulted clearly distinct from reference cultivars in multivariate analysis. In general, no particular structure was found in old cultivars, but a strong differentiation of ‘Agridulce’ and ‘Hojancas’ is reported due to their bigger fruits. Our results support the molecular analysis and call for further analysis of the local apple germplasm and long-term conservation actions.
Keywords
Apple genetic resources, old apple cultivars, phenotype, multivariate analysis
Introduction
The apple tree (Malus domestica Borkh.) is the most important temperate fruit tree crop, with more than 126 million tonnes harvested worldwide in 2020 (FAO, 2022). Such production is in line with the Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture (FAO, 2010), which reported Malus L. genetic resources to be among the largest ex situ collections. Morphological characterization of Malus has been essential for an adequate description of germplasm collections, for breeding programmes (Božović, Lazović, Ercisli, Adakalić, & Jaćimović, 2015) and taxonomic studies (Höfer, Mohamed, Eldin, Sellmann, & Peil, 2014; Wagner, Maurer, Lemmer, Schmitt, & Wagner, 2014). Currently, although the information provided by genetic markers (such as microsatellites) is preferred against phenotyping due to their stability and economy (Ban, Yun, Kim, Kwon, & Choi, 2014; Reddy, Sarla, & Siddiq, 2002), the study of agricultural germplasm by morphological traits is still relevant and useful in diversity analysis (Božović et al., 2015; Király, Ladányi, Nagyistván, & Tóth, 2015; Kumar, Singh, Pramanick, Verma, & Srivastav, 2018).
As a result, apple morphological descriptions were conducted in Bosnia and Herzegovina (Gaši, Šimon, Pojskić, Kurtović, & Pejić, 2011), Canada (Watts, Migicovsky, & Mcclure, 2021), Hungary (Király et al., 2015), India (Dolker, Kumar, Chandel, Angmo, & Chaurasia, 2021), Iran (Farrokhi, Darvishzadeh, Maleki, Naseri, & L, 2013), Italy (Martinelli, Busconi, Camangi, Fogher, & Stefani, 2008), the Kashmir Valley (Dar, Wani, & Dhar, 2015), Macedonia (Kiprijanovski, Arsov, & Saraginovski, 2020), Montenegro (Božović et al., 2015), Serbia (Mratinić, Akšić, & M, 2012) and Turkey (Karatas, 2022), reporting high morphological diversity. The most common morphological descriptors used in those works belong to international guidelines such as IBPGR (1982) and (UPOV, 2005) and they focus on fruit characteristics because sensorial characteristics and consumer demand focus on fruits (Pereira-Lorenzo, Fischer, Ramos-Cabrer, & Castro, 2018).
In Spain, several studies also reported great phenotypic apple diversity (Pérez-Romero, Suárez, Dapena, & Rallo, 2015; Ramos-Cabrer, Díaz-Hernández, & Pereira-Lorenzo, 2007; Royo & Itoiz, 2004; Santesteban, Miranda, & Royo, 2009), but they did not include old cultivars from some central regions. This lack of information should be filled, as some of these undescribed old apple cultivars are valued for their sensorial qualities, others are suspected to be exclusive to central Spain, and the abandonment of rural landscapes threatens their survival (Aceituno-Mata, 2010; Arnal, Gogorcena, Tardío, Roldán-Ruiz, & Lázaro, 2020; Comunidad de Madrid, 2018).
This study aims to quantify the morphological diversity of old apple cultivars growing in rural areas of central Spain, compare it with a previous genetic study with simple sequence repeats (Arnal et al., 2020) and see whether the old traditional cultivars were distinguishable by morphological descriptors. This work also provides the foundations for further agronomic and sensorial studies to complete the Spanish apple morphological description and provide valuable information that will increase the knowledge of apple genetic resources.
Material and Methods
Plant material
A collection of 67 individuals from 41 accessions belonging to 23 old Spanish apple cultivars was evaluated to assess their morphological diversity (Table 1). The collection is located in Arganda del Rey (Madrid) and belongs to the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario IMIDRA (Figure 1). For each accession, in 2009 two scions were grafted onto seedlings of M. domestica in a frame of 5m, being the aisles oriented in the SW-NE direction. Two individuals of nine main reference varieties and sports (grafted in rootstocks from the same nursery) curated in the same orchard and environmental conditions were included as controls: ‘Fuji Aztec’, ‘Fuji Kiku 8’, ‘Gala Buckeye’, ‘Gala Schniga’, ‘Golden Delicious’, ‘Golden Reinders’, ‘Granny Smith’, ‘Reineta Blanca’ and ‘Verde Doncella’ (Table 2). Those references were selected because they are widespread in Spain (Iglesias, Carbó, Bonany, & Montserrat, 2009).
The annual maintenance of the collection was conducted as follows: goblet pruning at the end of autumn, soil amendment at the beginning of winter and a preventive application of pesticides against aphids at the end of spring. Trees were irrigated every two weeks from May to September to reduce water stress during summer.
Morphological descriptors
A set of 67 morphological descriptors (25 quantitative and 42 qualitative, of which 3 were discrete, 23 nominal and 16 ordinal) were assessed on for 4 organs: 8 descriptors on winter 1-year-old wooden branches (or shoots), 15 on leaves, 16 on flowers and 28 on fruits (Table 3). The descriptors were obtained from IBPGR (1982), (UPOV, 2005) and Urbina and Dalmases (2014) and new descriptors and further categories in some qualitative traits were also considered, such as watercore (Arnal, 2021). Ten (10) to 20 fruits, 20 leaves, 10 flowers and 20 shoots were collected from different orientations of the tree crown in two years (except flowers) from summer 2016 to autumn 2019 and stored at 4–7ºC until processing. In particular, leaves and shoots were collected in 2016 and 2017, and flowers and fruits through the four years of the study.
Continuous descriptors of shoots, flowers and fruits were measured manually using a JP Selecta model 5900601 digital caliper with a precision of 0.01mm. Leaf quantitative descriptors were captured with ImageJ (Schneider, Rasband, & Eliceiri, 2012), so leaves were previously scanned attached to a 2D-scale. Finally, apple fruit weight (ten per individual) was registered with a Sartorius CP 2202 S digital scale with a precision of 0.01g.
Data analysis
Phenotypic diversity. Arithmetic means for 25 quantitative, medians for 16 discrete and ordinal, and modes for 26 nominal descriptors were calculated to obtain the central values by accession and cultivar. All qualitative descriptors were translated into a numerical value to meet computing requirements. Student’s t- and Cohen’s d-tests were conducted to identify ifferences between types of cultivar descriptors (references and old apple cultivars). Lastly, Tukey’s HSD test was performed to detect different groups among old apple cultivars. The significance level (α) was set at 0.05.
Correlations. A correlation matrix was calculated to explore significant correlations between descriptors in old traditional cultivars. Correlations between continuous descriptors were done with Pearson, whereas the rest were computed with Spearman. No correlations between nominal descriptors were performed. Descriptors with no variance were removed at this stage.

|
Old cultivar name |
Individual/s |
Accession |
Collection site |
|
Amarillo de El Paular |
APRA1/2 |
APRA |
Rascafría |
|
Camuesa |
CABU1/2 |
CABU |
Bustarviejo |
|
CAPR1/2 |
CAPR |
Prádena del Rincón |
|
|
Camueso tardío |
CTAP1/2 |
CTAP |
Puebla de la Sierra |
|
Camueso temprano |
CTEP1 |
CTEP |
Puebla de la Sierra |
|
de Chapa |
CHCA1/2 |
CHCA |
Canencia |
|
CHVA1 |
CHVA |
Valdemanco |
|
|
Esperiega |
ESPU1/2 |
ESPU |
Puebla de la Sierra |
|
Agridulce |
MAMO1/2 |
MAMO |
Montejo de la Sierra |
|
Hojancas |
MHHO1/2 |
MHHO |
Horcajuelo de la Sierra |
|
MHPR1/2 |
MHPR |
Prádena del Rincón |
|
|
del Ortel |
ORCA1/2 |
ORCA |
Canencia |
|
ORMO3 |
ORMO |
Morata de Tajuña |
|
|
Pero de Aragón |
PAHO1/2 |
PAHO |
Horcajuelo de la Sierra |
|
PAPR1/2 |
PAPR |
Prádena del Rincón |
|
|
PAPU1/2 |
PAPU |
Puebla de la Sierra |
|
|
Pepita de melón |
PECA1/2 |
PECA |
Canencia |
|
PEHI1/2 |
PEHI |
La Hiruela |
|
|
PEVA2 |
PEVA |
Valdemanco |
|
|
Pero gordo |
PGHI2 |
PGHI |
La Hiruela |
|
Pero pardo |
PPHI1 |
PPHI |
La Hiruela |
|
PPMO1/2 |
PPMO |
Montejo de la Sierra |
|
|
PPPU1/2 |
PPPU |
Puebla de la Sierra |
|
|
Pero real |
PRBU1/2 |
PRBU |
Bustarviejo |
|
PRHI1/2 |
PRHI |
La Hiruela |
|
|
Rabudas |
RAHI1/2 |
RAHI |
La Hiruela |
|
Reineta |
REHO11 |
REHO1 |
Horcajuelo de la Sierra |
|
REHO21 |
REHO2 |
||
|
REMO1 |
REMO |
Montejo de la Sierra |
|
|
REPR1/2 |
REPR |
Prádena del Rincón |
|
|
Rojillo |
RJHO1/2 |
RJHO |
Horcajuelo de la Sierra |
|
Rojo |
RJPR1/2 |
RJPR |
Prádena del Rincón |
|
RJPU1/2 |
RJPU |
Puebla de la Sierra |
|
|
de Rosa |
RORA1/2 |
RORA |
Rascafría |
|
Rojillo temprano |
RTEV1 |
RTEV |
Valdemanco |
|
San Felipe |
SFCA1 |
SFCA |
Carabaña |
|
Temprano |
TEPI1 |
TEPI |
Pinilla del Valle |
|
Verde Doncella |
VDCA1/2 |
VDCA |
Canencia |
|
VDHO1/2 |
VDHO |
Horcajuelo de la Sierra |
|
|
VDTI1 |
VDTI |
Tielmes |
|
Reference cultivar name |
Accession number |
Origin |
|
Fuji Aztec |
4 |
Worldwide cultivar |
|
7 |
||
|
Fuji Kiku 8 |
11 |
Worldwide cultivar |
|
14 |
||
|
Gala Buckeye |
115 |
Worldwide cultivar |
|
117 |
||
|
Gala Schniga |
109 |
Worldwide cultivar |
|
113 |
||
|
Golden Delicious |
104 |
Worldwide cultivar |
|
108 |
||
|
Golden Reinders |
101 |
Worldwide cultivar |
|
203 |
||
|
Granny Smith |
206 |
Worldwide cultivar |
|
210 |
||
|
Reineta Blanca |
211 |
Worldwide cultivar |
|
215 |
||
|
Verde Doncella |
307 |
National cultivar |
|
311 |
Multivariate analysis. A principal coordinate analysis (PCoA) was performed to visualize the possible groups of cultivars and detect the descriptors that better describe the differences among individuals. In the cluster analysis, a distance matrix between accessions was calculated with Nei’s distance (Nei, 1973) and the dendrogram was plotted using the unweighted pair group method with arithmetic mean (UPGMA) hierarchical agglomerative method (Sokal & Michener, 1958).
Analysis computing. Statistics were performed in R Studio v.3.4.1 (R Core Team, 2017) and a set of packages: ‘effsize’ for Cohen’s d-test (Torchiano, 2018), ‘corrplot’ (Wei & Simko, 2017) for the correlation matrix and ‘agricolae’ (Mendiburu, 2019) for Tukey HSD. Multivariate analysis was computed with an adapted version of the ‘MorphoTools’ script (Koutecký, 2015).
Results
Phenotypic diversity
Thirty-two morphological descriptors (48% of the total; 19 continuous and 13 non-continuous), showed significant differences between references and old apple cultivars (Table 4). In general, old apple cultivars registered lower quantitative values than references, but level frequencies in many qualitative descriptors were less skewed.
Shoots, leaves and flowers from both types of cultivars were similar, as there were only 14 significant descriptors out of 39 (36%). Some significantly different descriptors found in those organs were the width of the apical bud (ShW; P = 6.63 × 10-14), the petal length (FlPetL; P = 0.0012) and the petal width (FlPetW; P = 0.023).
In fruit, 18 out of 28 descriptors were significantly different (64%). The calyx opening diameter (FrCCD; P = 4.26 × 10-24), fruit peduncle length (FrPedL; P = 5.57 × 10-37), and peduncle width (FrWP; P = 7.37 × 10-18) stood out by their significance and effect size (Cohen’s d). In fact, it was observed that the peduncles from old apple cultivars were around 1cm shorter than those from references (Figure 2a). Other important quantitative descriptors such as the peduncular cavity width (FrSCW; P = 0.0163), the calyx cavity width (FrCCW; P = 0.0163), fruit length (FrL; P = 0.0026) and fruit weight (FrM; P = 0.0275) were significant, but the effect size (Cohen’s d) was not large. Alternatively, no significant differences existed in fruit width (FrW; P = 0.2081). In the fruit qualitative descriptors, the depth of the calyx cavity (FrCCDep) was 'intermediate' in old apple cultivars and 'strong' in references, with significant differences (P = 0.006), describing more diversity in old apple cultivars, as it was relatively easy to find apples with a 'weak', 'intermediate', 'strong', or 'very strong' calyx. Regarding over colour (FrUpCol), it was found that old apple cultivars had significantly less over colour than references (P = 2.69 × 10-06). Nevertheless, among apples with cheeks, 'red' and 'yellow' were the most abundant colours.
Means, medians and modes were also computed by cultivar. As a result, means of quantitative descriptors from references were contained in the Tukey’s HSD groups of old apple cultivars. The average fruit length (FrL) of the apples was in the 43–63mm range and the fruit width (FrW) was between 50 and 81.5mm. Regarding fruit weight (FrM), apples weighed 125g on average. Their shape (FrShp) was mostly conical (sum of 'conical globose', 'conical oblong' and 'conical truncated'), with a minority of ellipsoidal and flat globose shapes.
The cultivar ‘Agridulce’ showed significant larger sizes than the other old apple cultivars and even references, as its measurements belonged to the 'a' group of Tukey’s HSD in 17 out of the 25 quantitative descriptors (almost 50%), such as fruit length (FrL), fruit weight (FrM) and fruit width (FrW). ‘Hojancas’ and ‘Pero gordo’ also tended to have larger organs. The rest of the cultivars presented intermediate size organs, except ‘Esperiega’, and ‘San Felipe’, which showed small organs.
Correlations
There was significant correlation in 44 out of 67 descriptors studied, as the total average significant correlation was 0.43 (Figure 3). Shoot colour (ShCol) was removed as no variance was detected. By organ, the means of correlation was 0.37 in the shoot, 0.54 in the leaf, 0.42 in the flower, and 0.44 in the fruit. Correlations within each organ were mainly positive, such as leaf area
|
Organ |
Descriptor name |
Code |
Type |
Levels |
Source |
|
Shoot |
Pubescence on the apical bud |
ShBPub |
O |
0, glabrous; 1, intermediate; 2, tomentose |
3 |
|
Shoot colour |
ShCol |
1, brown; 2, reddish brown, 3; green; 4, grey; 5, purple; 6, red; 7, brown reddish; 8, light brown |
2 |
||
|
|
Shoot diameter (mm) |
ShDia |
C |
-- |
2 |
|
|
Length of the apical bud (mm) |
ShL |
C |
-- |
3 |
|
|
Lenticels# |
ShLent |
O |
1, very few; 3, few; 5 frequent; 7, densely populated |
2, 3 |
|
|
Apical shoot shape# |
ShShp |
1, semispherical; 2, ovoid; 3 intermediate; 4, conical |
3 |
|
|
|
Pubescence on shoot |
ShSPub |
O |
0, glabrous; right, 9, tomentose |
2 |
|
Width of the apical bud (mm) |
ShW |
C |
-- |
4 |
|
|
Leaf |
Leaf area (cm2) |
LeArea |
C |
-- |
4 |
|
|
Asymmetry of the leaf blade |
LeAsim |
0, symmetric; 1, asymmetric |
4 |
|
|
|
Shape of the base of the leaf blade |
LeBas |
1, cuneate; 2, rounded cuneate; 3, rounded; 4, asymmetric; 5, cordate; 7, truncated |
3 |
|
|
|
Petiole colour |
LeCol |
1, purple; 2, green and purple; 3, green |
2 |
|
|
|
Leaf edge shape |
LeEdg |
1, crenate; 2, bicrenate; 3, serrate-1; 4, serrate-2; 5, biserrate-2; 6, biserrate-1; 7, triserrate |
2, 3 |
|
|
Foliar blade folding |
LeFold |
1, folded; 2, turned; 3, convex; 4, ondulate; 5, flat |
3 |
||
|
Leaf blade length (cm) |
LeL |
C |
-- |
2, 3 |
|
|
Maximum width of the leaf blade (cm) |
LeMWL |
C |
-- |
4 |
|
|
Petiole length (cm) |
LePetL |
C |
-- |
3 |
|
|
Pubescence on the reverse |
LePub |
O |
0, not pubescent; 1, pubescent at the base of the midrib; 3, little pubescent; 5, pubescent; 7, very pubescent; 9, tomentose |
2, 3 |
|
|
Leaf blade shape |
LeShp |
O |
1, ovate; 2, elliptical; 3, obovate |
4 |
|
|
Leaf petiole stipules |
LeSti |
1, rudimentary; 3, short filiform; 5, long filiform; 7, narrow foliar; 9, wide foliar |
3 |
||
|
Leaf apex length (mm) |
LeTip |
C |
-- |
3 |
|
|
Shape of the leaf apex |
LeTipShp |
2, rounded; 3, acute; 5, mucronate; 7, acuminated; 9, cuspidate |
4 |
||
|
Leaf blade width (cm) |
LeW |
C |
-- |
2, 3 |
|
|
Flower |
Androecium length (mm) |
FlAnd |
C |
-- |
4 |
|
Dominance of the gynoecium over androecium |
FlDom |
O |
1, dominated; 2, balanced; 3, dominant |
2 |
|
|
|
Gynoecium length (mm) |
FlGin |
C |
-- |
3 |
|
|
Pedicel colour |
FlPedCol |
O |
1; purple; 2, purple and green; 3, green |
4 |
|
|
Flower pedicel length (mm) |
FlPedL |
C |
-- |
4 |
|
|
Petal colour |
FlPetCol |
0, white; 1, pink white; 2, purple white; 3, purple |
3 |
|
|
|
Petal length (mm) |
FlPetL |
C |
-- |
2, 3 |
|
|
Number of petals |
FlPetN |
D |
Integer counting |
3 |
|
|
Petal width (mm) |
FlPetW |
C |
-- |
2, 3 |
|
|
Pubescence on pedicel |
FlPub |
O |
1, glabrous; 2, slightly pubescent; 3, tomentose |
4 |
|
Relative position of the petals |
FlRPP |
O |
0, free; 1, tangent; 2, overlapped |
2, 3 |
|
|
Sepal length (mm) |
FlSepL |
C |
-- |
4 |
|
|
Flower shape |
FlShp |
1, flat turned; 2, turned cupuliform; 3, cupuliform; 4, slightly cupuliform; 5, flat |
3 |
||
|
Flower pedicel stipules |
FlSti |
0, no present; 1, present |
4 |
||
|
Type of petal |
FlTyp |
0, flat; 1, wavy; 2, concave; 3, convex |
4 |
||
|
|
Welding point of the stamens |
FlWeld |
O |
1, welded at the base; 2 welded in pairs up to a certain height; 3 completely welded |
3 |
|
Fruit |
Calyx opening diameter (mm) |
FrCCD |
C |
-- |
2, 3 |
|
Depth of the calyx cavity# |
FrCCDep |
O |
0, external; 1, very weak; 2, weak 3, intermediate; 4, strong; 5, very strong |
2, 3 |
|
|
|
Length of the calyx cavity (mm) |
FrCCL |
C |
-- |
2, 3 |
|
|
Shape of the opening of the calyx cavity |
FrCCShp |
0, without sepals; 1, convergent; 2, partially extended or extended; 3, erect |
2, 3 |
|
|
|
Calyx cavity width (mm) |
FrCCW |
C |
-- |
2, 3 |
|
|
Over colour distribution# |
FrDisCol |
O |
0, uniform (no cheeks); 1, blurred; 2, blurred and stripped; 3, stripped |
1, 3 |
|
|
Opening of the calyx cavity# |
FrEye |
0, closed; 1 open |
3 |
|
|
|
Flattening# |
FrFlat |
O |
1, dominated; 2, balanced; 3 dominant |
2, 3 |
|
|
Surface colour |
FrGroCol |
1, green; 2, light green; 3, yellowish green; 4, light yellow and 5, yellow |
1, 2, 3 |
|
|
|
Opening of the locules |
FrHea |
O |
0, closed; 1, semi-open; 2 open |
2, 3 |
|
|
Fruit length (mm) |
FrL |
C |
-- |
2 |
|
|
Number of loculi |
FrLoc |
D |
Integer counting |
1 |
|
|
Fruit weight (g) |
FrM |
C |
-- |
1, 3 |
|
|
Pulp colour |
FrMCol |
0, white; 1, greenish white; 2, green; 3, yellowish green; 4, white-yellowish; 5, Greenish yellow; 6, yellow |
2, 3 |
|
|
|
Fruit peduncle length (mm) |
FrPedL |
C |
-- |
2, 3 |
|
|
Russeting in the calyx cavity# |
FrRCC |
0, no russeting; 1, russeting |
2, 3 |
|
|
|
Russeting on fruit faces# |
FrRF |
0, no russeting; 1, russeting |
2, 3 |
|
|
Ribs |
FrRib |
O |
from 1, absent; to 5, very prominent |
2, 3 |
|
|
Russeting in the peduncular cavity# |
FrRS |
0, no russeting; 1, russeting |
2, 3 |
||
|
|
Peduncular cavity length (mm) |
FrSCL |
C |
-- |
2, 3 |
|
|
Peduncular cavity width (mm) |
FrSCW |
C |
-- |
2, 3 |
|
|
Number of seeds |
FrSeed |
D |
Integer counting |
4 |
|
Fruit shape |
FrShp |
1, Globose 2, conical globose; 3, wide conical globose; 4, flat; 5, flat globose; 6, conical; 7, narrow conical; 8, conical truncate; 9 ellipsoidal; 10, conical ellipsoidal; 11, oblong; 12, conical oblong; 13, asymmetric |
1, 2, 3 |
||
|
Over colour |
FrUpCol |
0, without over colour (no cheeks); 1, white; 2, yellowish; 3, yellow; 4, orange; 5, reddish-pink; 6, red; 7, purple; 9, brown |
1, 2, 3 |
||
|
Vitrification (or watercore) |
FrVitr |
0, absent; 1, present |
4 |
||
|
Fruit width (mm) |
FrW |
C |
-- |
2 |
|
|
|
Bloom of skin |
FrWax |
O |
1; weak; 2, moderate; 3, intense |
2, 3 |
|
Peduncle width (mm) |
FrWP |
C |
-- |
2, 3 |
|
Organ |
Descriptor |
Type |
Mean |
Median |
Mode |
P-value |
Sig. |
Cohen's d |
|||
|
Ref. |
OAC |
Ref. |
OAC |
Ref. |
OAC |
||||||
|
Shoot |
ShCol |
N |
|
|
|
|
Brown |
2.69 × 10-5 |
*** |
- |
|
|
ShDia |
C |
4.95±0.06 |
6.81±0.07 |
|
|
|
|
8.74 × 10-94 |
*** |
Large |
|
|
ShL |
C |
6.75±0.08 |
5.41±0.05 |
|
|
|
|
1.74 × 10-41 |
*** |
Large |
|
|
ShW |
C |
4.54±0.05 |
4.13±0.04 |
|
|
|
|
6.63 × 10-14 |
*** |
Medium |
|
|
Leaf |
LeFold |
N |
|
|
|
|
Folded |
0.003 |
** |
- |
|
|
LeMWL |
C |
4.48±0.17 |
4.72±0.07 |
|
|
|
|
0.013 |
* |
Small |
|
|
LeTip |
C |
0.62±0.03 |
0.68±0.01 |
|
|
|
|
0.006 |
** |
Small |
|
|
Flower |
FlAnd |
C |
10.46±0.53 |
9.96±0.24 |
|
|
|
|
0.001 |
** |
Small |
|
FlPedL |
C |
17.43±0.93 |
8.66±0.25 |
|
|
|
|
2.20 × 10-16 |
*** |
Large |
|
|
FlPetL |
C |
23.28±1.18 |
21.85±0.52 |
|
|
|
|
0.001 |
** |
Small |
|
|
FlPetN |
D |
|
|
Five |
|
|
0 |
*** |
- |
||
|
FlPetW |
C |
16.27±0.82 |
15.58±0.38 |
|
|
|
|
0.023 |
* |
Small |
|
|
FlSepL |
C |
7.54±0.39 |
7.01±0.17 |
|
|
|
|
0.014 |
* |
Small |
|
|
FlShp |
N |
|
|
|
|
Flat |
0.034 |
* |
- |
||
|
Fruit |
FrCCD |
C |
5.88±0.22 |
3.81±0.13 |
|
|
|
|
4.26 × 10-24 |
*** |
Large |
|
FrCCDep |
O |
|
|
Strong |
Intermediate |
|
|
0.006 |
** |
- |
|
|
FrCCL |
C |
11.93±0.38 |
10.44±0.26 |
|
|
|
|
3.47 × 10-10 |
*** |
Medium |
|
|
FrCCW |
C |
24.82±0.76 |
24±0.57 |
|
|
|
|
0.016 |
* |
Negligible |
|
|
FrDisCol |
O |
|
|
Blurred |
Uniform |
|
|
0.012 |
* |
- |
|
|
FrGroCol |
N |
|
|
|
|
Yellow |
2.46 × 10-10 |
*** |
- |
||
|
FrL |
C |
57.35±1.71 |
55.57±1.29 |
|
|
|
|
0.003 |
** |
Small |
|
|
FrLoc |
D |
|
|
Five |
|
|
2.20 × 10-16 |
*** |
- |
||
|
FrM |
C |
131.87±4.47 |
124.37±3.27 |
|
|
|
|
0.028 |
* |
Negligible |
|
|
FrMCol |
N |
|
|
|
|
Greenish-yellow |
6.12 × 10-05 |
*** |
- |
||
|
FrPedL |
C |
24.36±1.01 |
12.54±0.38 |
|
|
|
|
5.57 × 10-37 |
*** |
Large |
|
|
FrRS |
N |
|
|
|
|
Presence |
0.017 |
* |
- |
||
|
FrSCL |
C |
2.42±0.47 |
3.64±0.29 |
|
|
|
|
7.67 × 10-20 |
*** |
Medium |
|
|
FrSCW |
C |
29.71±0.91 |
27.94±0.67 |
|
|
|
|
2.64 × 10-05 |
*** |
Small |
|
|
FrUpCol |
N |
|
|
|
|
Absent |
2.69 × 10-06 |
*** |
- |
||
|
FrVitr |
N |
|
|
|
|
Absent |
- |
*** |
- |
||
|
FrWax |
O |
|
|
Very intense |
|
|
0.011 |
* |
- |
||
|
FrWP |
C |
2.65±0.10 |
3.42±0.09 |
|
|
|
|
7.37 × 10-18 |
*** |
Large |
|


(LeArea) with leaf width (LeW; 0.91), petal length (FlPetL) with petal width (FlPetW; 0.81) and fruit weight (FrM) with fruit width (FrW; 0.92) and fruit length (FrL; 0.71).
Some significant correlations were negative, such as the one found between the fruit peduncle length (FrPedL) and peduncle width (FrWP; -0.68, Figure 2b), and the length of the apical bud (ShL) with peduncle width (FrWP; -0.63). Correlations between two different organs were also positive, for example between the fruit peduncle length (FrPedL) and flower pedicel length (FlPedL; 0.86) and with length of the apical bud (ShL; 0.72). We found some negative significant correlations, such as the ones that involved the flower pedicel length (FlPedL) with peduncle width (FrWP; -0.63) and with the length of the apical bud (ShL; -0.55).
Multivariate analysis
The two multivariate analyses showed that the morphological diversity of apple references was lower than the one from old cultivars. Results from PCoA (Table 5) showed that the first three PCos explained 31% of the variability assessed, with seven coordinates necessary to reach 50% variability. By coordinates, PCo 1 represented 14% of the variance, PCo 2 10%, and PCo 3 7%. The most important correlations in PCo 1 were negative: outstanding fruit weight (FrM; -0.76), fruit length (FrL; -0.75), leaf area (LeArea; -0.75), peduncular cavity width (FrSCW;-0.74) and leaf width (LeW; -0.71). In PCo 2, the balance between negative and positive correlations was similar, with important correlations for peduncle width (FrWP; -0.77), peduncular cavity width (FrSCW; -0.740), leaf width (LeW; -0.71), length of the apical bud (ShL; 0.70), flower pedicel length (FlPedL; 0.67) and lenticels (ShLent; 0.62). In PCo 3, the strongest positive correlations involved the maximum width of the leaf blade (LeMWL; 0.65), leaf length (LeL; 0.54), and apical shoot shape (ShShp; 0.49). Among the negative correlations in PCo 3, most noticeable were the width of the apical bud (ShW; -0.51) and over colour (FrUpCol; -0.48). In the plot that represents apple individuals in PCo 1 and PCo 2 (24% of variance), we detected that clonal replicates of references were closely grouped while old apple cultivars appeared separated from apple references, but no further structure was detected in the traditional pool (Figure 4). Despite their low structure in the plot, ‘Agridulce’ (MAMO1 and MAMO2) and ‘Hojancas’ (MHHO1) were clearly distinct from the rest of the cultivars thanks to quantitative descriptors such as leaf area (LeArea), fruit weight (FrM) and fruit width (FrW). Finally, ‘Verde Doncella’ with VDCA, VDHO and VDTI clustered together.
|
Descriptor |
PCo 1 |
PCo 2 |
PCo 3 |
|
Descriptor |
PCo 1 |
PCo 2 |
PCo 3 |
|---|---|---|---|---|---|---|---|---|
|
FlAnd |
-0.59 |
0.08 |
-0.04 |
|
FrRib |
0.12 |
-0.09 |
-0.15 |
|
FlDom |
-0.26 |
-0.45 |
-0.06 |
|
FrRS |
0.03 |
-0.21 |
-0.21 |
|
FlGin |
-0.66 |
-0.34 |
-0.03 |
|
FrSCL |
-0.68 |
0.45 |
0.07 |
|
FlPedCol |
-0.13 |
0.39 |
-0.22 |
|
FrSCW |
-0.74 |
-0.26 |
-0.08 |
|
FlPedL |
-0.61 |
0.51 |
-0.30 |
|
FrSeed |
0.10 |
0.55 |
-0.14 |
|
FlPetCol |
-0.13 |
0.23 |
0.14 |
|
FrShp |
-0.26 |
0.53 |
0.25 |
|
FlPetL |
-0.60 |
-0.01 |
0.06 |
|
FrUpCol |
0.08 |
0.43 |
-0.48 |
|
FlPetN |
0.24 |
-0.15 |
0.12 |
|
FrVitr |
0.22 |
-0.12 |
0.19 |
|
FlPetW |
-0.69 |
-0.22 |
-0.05 |
|
FrW |
-0.70 |
-0.41 |
-0.10 |
|
FlPub |
-0.01 |
0.07 |
0.21 |
|
FrWax |
-0.14 |
-0.12 |
0.22 |
|
FlRPP |
-0.12 |
-0.34 |
-0.07 |
|
FrWP |
0.11 |
-0.77 |
-0.03 |
|
FlSepL |
-0.68 |
0.04 |
0.14 |
|
LeArea |
-0.75 |
-0.19 |
0.29 |
|
FlShp |
-0.13 |
-0.05 |
0.38 |
|
LeAsim |
-0.03 |
0.10 |
0.24 |
|
FlSti |
-0.30 |
-0.03 |
-0.30 |
|
LeBas |
-0.18 |
-0.44 |
-0.29 |
|
FlTyp |
-0.16 |
-0.14 |
-0.23 |
|
LeCol |
-0.22 |
-0.26 |
0.15 |
|
FlWeld |
0.04 |
0.07 |
0.29 |
|
LeEdg |
-0.40 |
-0.09 |
0.24 |
|
FrCCD |
-0.55 |
0.07 |
-0.44 |
|
LeFold |
-0.09 |
-0.06 |
-0.004 |
|
FrCCDep |
-0.15 |
0.18 |
0.20 |
|
LeL |
-0.54 |
0.09 |
0.54 |
|
FrCCL |
-0.63 |
0.12 |
-0.05 |
|
LeMWL |
-0.43 |
0.15 |
0.65 |
|
FrCCShp |
-0.18 |
-0.02 |
0.10 |
|
LePetL |
-0.04 |
0.24 |
0.38 |
|
FrCCW |
-0.52 |
-0.31 |
-0.15 |
|
LePub |
0.19 |
-0.39 |
-0.16 |
|
FrDisCol |
0.10 |
0.37 |
-0.37 |
|
LeShp |
-0.03 |
0.21 |
0.44 |
|
FrEye |
0.08 |
-0.04 |
-0.29 |
|
LeSti |
-0.49 |
-0.08 |
-0.47 |
|
FrFlat |
0.12 |
-0.64 |
-0.36 |
|
LeTip |
-0.13 |
-0.22 |
0.17 |
|
FrGroCol |
0.02 |
0.18 |
0.40 |
|
LeTipShp |
-0.17 |
-0.21 |
-0.20 |
|
FrHea |
-0.26 |
-0.25 |
-0.13 |
|
LeW |
-0.71 |
-0.31 |
0.09 |
|
FrL |
-0.75 |
0.06 |
0.17 |
|
ShCol |
0.29 |
-0.05 |
0.18 |
|
FrLoc |
-0.07 |
-0.07 |
0.16 |
|
ShDia |
0.10 |
-0.59 |
0.36 |
|
FrM |
-0.76 |
-0.33 |
-0.03 |
|
ShL |
-0.29 |
0.70 |
-0.30 |
|
FrMCol |
-0.20 |
0.46 |
0.09 |
|
ShLent |
-0.23 |
0.62 |
-0.19 |
|
FrPedL |
-0.57 |
0.67 |
-0.14 |
|
ShShp |
0.03 |
-0.13 |
0.49 |
|
FrRCC |
-0.21 |
-0.35 |
-0.24 |
|
ShSPub |
0.16 |
0.19 |
0.01 |
|
FrRF |
0.03 |
0.07 |
-0.14 |
|
ShW |
-0.12 |
-0.002 |
-0.51 |

In the cluster analysis, the two ‘Agridulce’ individuals (MAMO1 and MAMO2) split off very early from the rest of the individuals (Figure 5). They were followed by a ‘Rojillo temprano’ (RTEV1), two ‘Pero de Aragón’ (PAPR2 and PAPU1), a ‘Camueso tardío’ (CTEP1), a ‘Pero gordo’ (PGHI2), and a ‘Temprano’ (TEPI1). The rest of the dendrogram was structured in three clusters. The first cluster was composed of two 'peros' (PPPU2 and PRBU2), two 'reineta' (‘Reineta Blanca’ and REPR2), ‘Hojancas’ (MHHO1), and one ‘Camueso’ (CAPR1). The second cluster contained 28 individuals, including the two reference ‘Verde Doncella’ (VDHO2). Here also appeared a pool consisting of the cultivar ‘Esperiega’ (ESPU1, ESPU2), ‘de Chapa’ (CHVA1, CHCA1, and CHCA2) and ‘Pepita de melón’ (PECA2, PEVA2), as well as a group that nested the cultivars ‘Rojo’ and ‘Rojillo’ (RJHO1, RJPU1, RJPU2, and RJPR2). This second cluster also included a ‘Camuesa’ (CABU2) and the three remaining individuals of ‘Pepita de melón’ (PEHI1, PEHI2, and PECA1). The third cluster was composed by references (‘Fuji’, ‘Gala’, ‘Golden’, and ‘Granny Smith’), as well as one ‘Reineta’ (REPR1) and two ‘Pero de Aragón’ (PAPR1 and PAPU2).

Discussion
Phenotypic diversity
The results of the present morphological analysis of shoots, leaves and flowers were similar to other morphological studies (Božović et al., 2015; Hassan, Bhat, Dar, Mir, & Pandith, 2017). Fruit size measures were highly variable, especially fruit length (FrL), fruit weight (FrM) and fruit width (FrW). Although averages of these three descriptors were similar to results reported by other works, the registered range was larger than those described in studies by (Mratinić, Akšić, & M, 2011), (Özrenk, Gündoğdu, Kaya, & Kan, 2011), (Király, Redeczki, Erdélyi, & Tóth, 2012),Božović et al. (2015), Pérez-Romero et al. (2015) and Posadas-Herrera, López, Gutiérrez-Rangel, Díaz-Cervantes, and Ibáñez-Martínez (2018).
According to (Pereira-Lorenzo, Ramos-Cabrer, Ascasíbar-Errasti, & Piñeiro-Andión, 2003), (Gaši et al., 2011),Božović et al. (2015) and Pérez-Romero et al. (2015), quantitative descriptors related to apple cavities (FrSCL, FrSCW, FrCCL, FrCCW) are informative because they are genetically controlled. Those four descriptors detected statistically significant differences among studied cultivars, but such significance may be due to their correlation to fruit length (FrL) and fruit width (FrW). In fact, the depth of the calyx cavity (FrCCDep), a qualitative descriptor that relativizes the calyx cavity width and the calyx cavity length with the global size of the fruit, showed an intermediate diversity, since the depth of this cavity was found to be 'intermediate' in many of our studied old apple cultivars, similarly to Božović et al. (2015). Furthermore, no 'external' cavity was found neither in our collections nor in Božović et al. (2015), being ‘Sisa’ the unique old apple cultivar reported with such characteristic (Zovko, Vego, & Zovko, 2010).
Related to fruit shape, our results agree with Božović et al. (2015), since their predominant shapes were 'conical' and 'obloid', with some presence of 'ellipsoidal' and 'globose'. Nevertheless, not all studies reported conical shapes as dominant, as Pırlak, Güleryüz, Aslantaş, and Eşitken (2003) found that the 'flat', 'conical' and 'spherical' shapes were all abundant. Similarly, Hassan et al. (2017) reported predominantly the shapes 'globose', 'obloid' and 'ellipsoid', finding only one conical old apple cultivar. Although a great morphological diversity in apple shapes is generally reported, probably some of these differences could be attributed to the high subjectivity of this descriptor (Currie, Ganeshanandam, Noiton, Garrick, & Shelbourne, 2000).
Discrepancies in shape should have affected other descriptors such as flattening (FrFlat), but our results were similar to those reported in other collections (Božović et al., 2015; Salkić, Jovović, Salkić, & Salkić, 2017), leading us to consider that old apple cultivars are, in general, wider than longer. We also support this conclusion, as although fruit length (FrL) was larger in references, no significant differences between types of cultivars were found in fruit width (FrW).
Apple skin colour is supposed to be a distinctive trait in apple cultivars. The fruit ground colour (FrGroCol) ranged in our collection from 'greenish white', 'green', 'greenish yellow' to 'yellow', similarly to Božović et al. (2015), (Mišić, 2002) and Zovko et al. (2010). Regarding over colour (FrUpCol), apples herein described were mainly cheekless, as the most common level for this descriptor was 'absent'. Therefore, the studied old apple cultivars have a more uniform colour than those reported in the collection of Božović et al. (2015) and Šebek (2013). Concerning our cheeked apples, the most common colour was 'red', as in (Mratinić et al., 2012) and Božović et al. (2015).
Correlations
Many of the significant correlations computed were logical, supporting the botanical description of the apple tree (Aedo, Garmendia, Navarro, & C, 1998; Terpó, 1981) and agree with Ganopoulos, Tourvas, Xanthopoulou, Aravanopoulos, and Avramidou (2018) and Farrokhi et al. (2013), as the strongest correlations occurred among quantitative descriptors and in the same organ. Also, we obtained a strong correlation between leaf area and descriptors related to fruit size, as mentioned by Migicovsky, Li, Chitwood, and Myles (2018).
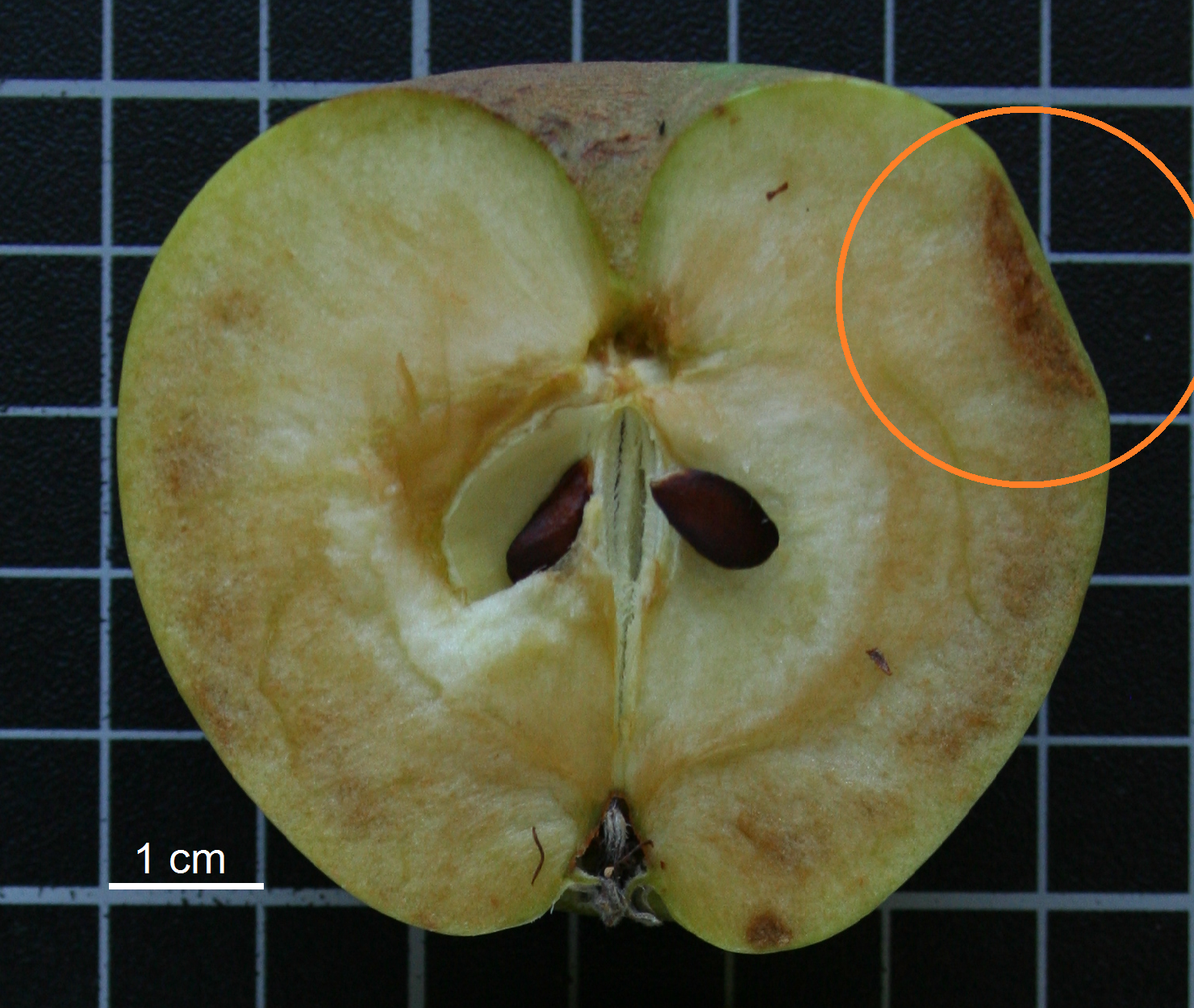
A significant correlation, important in breeding, was detected between the fruit peduncle length (FrPedL) and peduncle width (FrWP).Salkić et al. (2017) consider that short peduncles are not desirables. We agree with these authors, as probably short and wide peduncles may suffer from lack of growth space, causing some injuries to the fruits and decreasing their commercial quality (Figure 6). Although this behaviour was not deeply studied in this morphological characterization, our observations suggest that correlation analyses help breeders select descriptors that have a lever effect on genetic improvement (Chen & Lübberstedt, 2010; Ganopoulos et al., 2018).
Multivariate analysis
The PCoA decomposed the variance of the morphological descriptors analyzed. The sedimentation rate along the PCo is almost identical to the Spanish study of Pereira-Lorenzo et al. (2003), but it was slower compared to other collections (Božović et al., 2015; Ganopoulos et al., 2018; Gaši et al., 2011). For example, our PCo 1 only gathers 14% of the variance, whereas PCo 1 from Gaši et al. (2011) gathered almost 30% with 18 descriptors. A slow sedimentation rate does not necessarily indicate that our collection is more diverse than others. Probably, the greater the number of descriptors and accessions analyzed, the slower the sedimentation process tends to be. In fact, our study analyzed the largest number of descriptors, followed by Pereira-Lorenzo et al. (2003), who used 49 descriptors in 350 trees.
Regarding PCo eigenvalues, fruit descriptors were usually predominant, especially fruit weight and size. Our results were very similar to other studies, such as Božović et al. (2015), Gaši et al. (2011), Farrokhi et al. (2013) and Pereira-Lorenzo et al. (2003). Some results reported from other collections do not totally agree with ours. This is the case of Ganopoulos et al. (2018) where they highlighted other types of fruit characteristics, such as the number of loculi (FrLoc), pulp colour (FrMCol), russeting on fruit faces (FrRF) and calyx opening diameter (FrCCD). The importance of fruit descriptors in the total variance can be explained because it is the organ where selection is performed (Božović et al., 2015; Dar et al., 2015; Posadas-Herrera et al., 2018; Pérez-Romero et al., 2015; Salkić et al., 2017; Šebek, 2013).
Results from PCoA are consistent with the cluster analysis, as both detect a high morphological diversity in the old apple cultivars. Differences may be due to the lower number of references analyzed, as ‘Gala’, ‘Golden’, ‘Fuji’, and ‘Granny Smith’ are few, but they are the most widespread varieties in Spain and represent almost the whole apple production (Iglesias et al., 2009). In addition, we could discriminate references from old apple cultivars and we found an early separation of ‘Agridulce’, the presence of two clusters composed of ‘de Chapa’, ‘Esperiega’, ‘Pepita’, and ‘Camuesa’ and ‘Pepita’, respectively, and the inclusion of ‘Verde Doncella’ within old apple cultivars. The closeness of ‘Verde Doncella’ to other traditional apples is congruent with its breeding history, as this cultivar is autochthonous to Spain (Iglesias et al., 2009; Pina, Urrestarazu, & Errea, 2014; Urrestarazu, Miranda, Santesteban, & Royo, 2012).
We did not find further classification in old apple cultivars, as for instance we could not separate 'camuesas' from 'peros'. A clear separation of references from old traditional cultivars without a strong structure has been reported before (Božović et al., 2015; Ganopoulos et al., 2018), indicating that old apple germplasm is different from references, in contrast with Posadas-Herrera et al. (2018) and Király et al. (2015), who could not differentiate between both type of cultivars. Regarding descriptors, no single descriptor can distinguish among cultivars, but some of them may be informative and should be considered in cultivar classification, such as the flower pedicel length (FlPedL), depth of the calyx cavity (FrCCDep), fruit peduncle length (FrPedL) and peduncle width (FrWP).
Difficulties in old apple cultivar classification are probably due to boundaries among old apple cultivars being more diffuse than in modern cultivars, whose genealogy is always known (Laurens, 1999; Noiton & Alspach, 1996). For example, ‘Crisp Pink’ (Pink Lady®) derives genetically from ‘Golden’ and ‘Lady Williams’ (Iglesias et al., 2009). The lack of information on the origin of old apple cultivars is also accompanied by homonymies. Probably, any morphological or sensory trait may be enough to link two cultivars not necessarily parented. For example, flattened apples may evoke a 'reineta', as Martinelli et al. (2008) found that ‘Reineta grigia’ was not really a 'reineta'. Something similar was reported by Mratinić and Fotirić (2012), who informed that some accessions named ‘Šerbetka’ (which means 'too sweet') were later clustered separately.
Support of SSR molecular data in old apple cultivars identities
Morphological and DNA characterization are two complementary techniques, although conclusions about diversity and parental analysis are more robust with molecular analysis (Király et al., 2012). Therefore, we recently published a molecular analysis of the same accessions in this study based on 13 microsatellites (Arnal et al., 2020) in which we reported germplasm with breeding potential that should be further considered. Interestingly, PCoA and clustering analysis between the two studies are very comparable, as both clearly differentiate references from old apple cultivars and in general no further groups could be defined.
Our two studies pointed out two singular old apple cultivars (‘Agridulce’ and ‘Hojancas’), which may derive from 'reineta'. In the morphological study, these two old cultivars (especially ‘Agridulce’) showed traits that differentiated them better than microsatellites, as their differential morphology allowed us to segregate them even earlier than in the molecular study. In consequence, they could be considered for ex situ conservation and further studies. Moreover, both methodologies closely related ‘Camuesa’, ‘de Chapa’ and ‘Pepita’. Also, ‘Rojillo’ and ‘Rojo’, which seemed synonyms in our molecular analysis, showed similar morphological profiles, as they appeared together in the multivariate analysis. In contrast, the present morphological study does not gather all triploids in a cluster, nor detect the two groups of 'peros' found with microsatellites, as PGHI2, PAPU1, and PAPU2 fell each one in different clusters and one of them (PAPU2) was closely related to PAPR1.
In conclusion: 1) a great morphological diversity of old apple cultivars was detected in rural areas of central Spain; 2) the presented results confirm our previous analysis with microsatellites; 3) both approaches will help to better understand Spanish and global apple genetic resources; 4) the described collection contains two old apple cultivars (‘Agridulce’ and ‘Hojancas’) with a very distinct morphology, which may deserve further studies (such as flowering and ripening times, productivity, resistance to pests, etc.); 5) two old apple cultivars (‘de Chapa’ and ‘Pepita’) may be a variation of ‘Camuesa’, and 6) the cultivars ‘Rojillo’ and ‘Rojo’ are likely synonyms.
Funding
This study was supported by the research project FP16-ETNOB 'Prospection, characterization, ethnobotanical documentation and conservation of traditional horticultural varieties of the region of Madrid' funded by the local government of Madrid (Spain). The National Institute for Agricultural and Food Research and Technology (INIA) cofunded A. Arnal predoctoral contract.
Acknowledgements
A. Arnal thanks the National Institute for Agricultural and Food Research and Technology (INIA) for the predoctoral contract in agrifood, Yolanda Gogorcena for her comments on the draft, and Adrián Gómez for his editorial support.
Author contributions
A. Arnal, J. Tardío and A. Lázaro designed the research. A. Arnal sampled the individuals with the help of J. Tardío and A. Lázaro. A. Arnal adapted the R Code and analyzed the data. A. Arnal, J. Tardío and A. Lázaro wrote the manuscript. A. Arnal, J. Tardío and A. Lázaro revised the manuscript.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.